

常见多潜能标记物
多能性是细胞分化成任何细胞类型的能力。在个体中,诱导细胞多能性的能力可为开发性生物研究和疾病研究提供宝贵工具,并最终应用到个体化干细胞治疗中。Cell Signaling Technology (CST) 提供全系列的干细胞和细胞系标记物。我们的内部专家团队在相关研究应用中广泛验证了 CST™ 抗体,并且每种产品的验证数据可在线提供。请随时咨询我们的技术支持科学家,以防您需要熟练的合作伙伴。
关键的重编程因子: O—Oct4, K—Klf4, S—Sox2, M—c-Myc, E—Esrrb, Nr—Nr5a2, C—C/EBPα, L—Lin28, N—Nanog
| 物种 | 细胞类型 | 重编程因子 | 参考 |
|---|---|---|---|
| 小鼠 | 成纤维细胞 | OKSM, OKS, OSE, KSNr | Takahashi & Yamanaka 2006 |
| 成熟的 B 和 T 细胞 | OKSMC, OKSM | Hanna et al. 2008; Eminli et al. 2009 | |
| 髓样祖细胞 | OKSM | Eminli et al. 2009 | |
| 造血干细胞 | OKSM | Eminli et al. 2009 | |
| 脂肪源性干细胞 | OKSM | Sugii et al. 2010 | |
| 真皮乳头 | OKM, OK | Tsai et al. 2010 | |
| 卫星细胞 | OKSM | Tan et al. 2011 | |
| 胰腺 β 细胞 | OKSM | Stadtfeld et al. 2008 | |
| 肝内皮层 | OKS | Aoi et al. 2008 | |
| 神经干细胞 | OK | Kim et al. 2008; Kim et al. 2009 | |
| 黑色素细胞 | OKM | Utikal et al. 2009 | |
| 人 | 成纤维细胞 | OKSM, OSLN, OKS | Takahashi et al. 2007; Yu et al. 2007; Nakagawa et al. 2008 |
| 迁移的外周血 | OKSM | Loh et al. 2009 | |
| 脐带血内皮 | OSLN | Haase et al. 2009 | |
| 脐带血干细胞 | OKSM, OS | Eminli et al. 2009; Giorgetti et al. 2009 | |
| 脂肪源性干细胞 | OKSM, OKS | Sugii et al. 2010; Aoki et al. 2010 | |
| 肝细胞 | OKSM | Liu et al. 2010 | |
| 角质细胞 | OKSM, OKS | Aasen et al. 2008 | |
| 神经干细胞 | O | Kim et al. 2009 | |
| 胰腺 β 细胞 | OKSM | Bar-Nur et al. 2011 | |
| 羊膜细胞 | OKSM, OSN | Li et al. 2009; Zhao et al. 2010 |
表格改编自 Stadtfeld 和 Hochedlinger 2011。
对小鼠 iPS 细胞进行免疫荧光分析
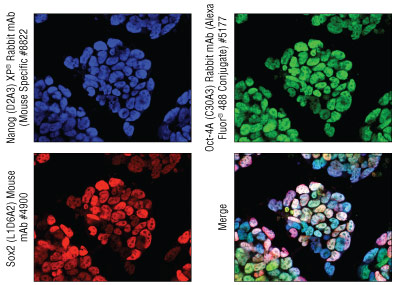
改编自 Walsh、Becker 和 Hochedlinger 2011。
正常发育期间在未分化的多能性胚胎干细胞和生殖细胞中表达的转录因子。连同 Sox2 和 Nanog 一起,它是维持多能电势所必需的。
发育期间在未分化的多能性胚胎干细胞和生殖细胞中表达的转录因子。连同 Oct-4 和 Nanog 一起,它是维持多能细胞电位所必需的。
含有同源结构域的转录因子,它是维持胚胎干细胞多能性和自我更新所必需的。Sox2 和关键多能性调节分子 Oct-4 等各种因子可调节表达。
含有锌指的转录因子 Krüppel 样因子 4 (KLF4);用于产生人和小鼠胚胎干 (ES) 细胞。
存在于人干细胞、畸胎癌细胞和胚胎生殖细胞的表面。
在小鼠胚胎癌细胞、胚胎干细胞和生殖细胞的表面表达的乳糖低聚糖,但仅在人生殖细胞中表达。分化时,SSEA1 在人细胞中的表达升高,而小鼠细胞分化则会导致表达下降。
在人畸胎癌干细胞、胚胎生殖细胞和胚胎干细胞的表面表达的糖脂碳水化合物。在人胚胎癌细胞分化后,人 SSEA4 的表达下降,但在小鼠细胞分化后,表达升高。