

了解 RNA 调节和表观转录组在疾病中的作用
简介
根据生物学中心法则,RNA 是将 DNA 中所含的基因组信息翻译成蛋白的分子。但这种简单的视图并不会触及 RNA 功能的复杂性。新试剂和技术(例如 RNA 交联和免疫沉淀法 (CLIP) 结合下一代测序)的开发使得全转录组分析变得可能,从而使得这个新兴领域的研究呈爆炸性增长。结果?关于 RNA 调节如何导致各种疾病进展的新见解。
转录后,信使 RNA (mRNA) 与 RNA 结合蛋白 (RBP) 相互作用,形成调节特定 mRNA 命运的信使核糖核蛋白 (mRNP) 复合体 (1)。这些包括加帽、剪切、运输、稳定性、沉默、衰变和翻译 (1, 2)。表观转录组是指 RNA 转录后修饰,包括腺苷和胞嘧啶甲基化、腺苷至肌苷编辑以及假尿苷化,它还在影响 mRNA 命运的过程中起到关键作用,并给基因表达调节又添加了一层复杂性。
RNA 甲基化
N6-甲基腺苷 (m6A)
N6-甲基腺苷 (m6A) 是最普遍且容易理解的 RNA 修饰,与组蛋白修饰类似,具有 m6A 特异性写入蛋白、读取蛋白和擦除蛋白。METTL3/METTL4 甲基转移酶复合体催化 m6A 与 YTHDC、YTHDF 和 eIF3 等 m6A 结合蛋白相互作用。这些相互作用介导 m6A 依赖性细胞事件,例如促进翻译和 RNA 稳定性。脱甲基酶(例如 FTO 和 ALKBH5)可逆转 m6A 修饰。证据表明 m6A 修饰在发育、T 细胞平衡以及胚胎干细胞的多能性中起到作用 (3)。
异常 m6A 修饰与各种癌症有关 (4)。写入蛋白、读取蛋白和擦除蛋白促进肿瘤发生所涉及的机制似乎特定于受影响的靶标 mRNA 以及癌症类型。例如,急性髓性白血病 (AML) 细胞中的 METTL3 水平升高会导致抗凋亡基因 BCL-2 过表达,但胶质母细胞瘤细胞中的 METTL3 水平降低会导致癌基因 ADAM19 表达增加。
证据表明 m6A 修饰还在多巴胺能神经元死亡(帕金森病 (PD) 的一个标志)中起到作用 (5)。在 PD 大鼠脑的纹状体中观察到 m6A 水平整体降低,并且 FTO 在多巴胺能细胞中过表达会诱导 NMDA 受体 1 表达,并使氧化应激和 CA2+ 水平升高,从而诱导凋亡。FTO 基因内含子 1 和内含子 2 的基因变体还与阿尔茨海默病 (AD) 有关。在欧洲血统和加勒比海西班牙裔白人中观察到这些地区有 SNP。与对照相比,AD 患者的 FTO 表达水平也较低 (6)。
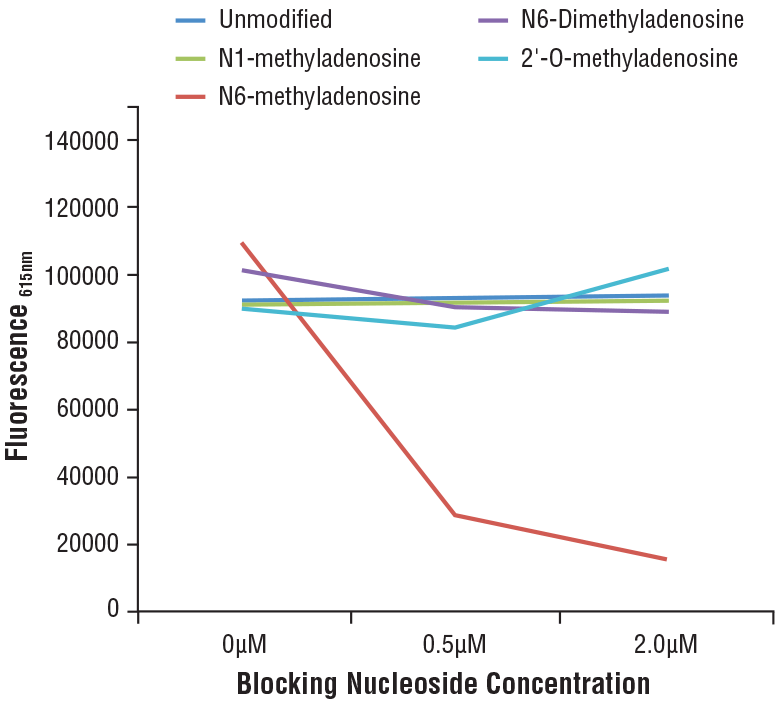
N6-Methyladenosine (m6A) (D9D9W) Rabbit mAb #56593:N6-Methyladenosine (m6A) (D9D9W) Rabbit mAb 的特异性通过竞争性 ELISA 进行确定。该图表描绘了在不同修饰腺苷浓度递增的情况下抗体与预包被 m6A 寡核苷酸的结合。如图表所示,抗体结合仅会被游离 m6A 核苷阻断。
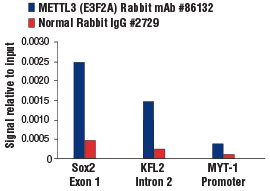
METTL3 (E3F2A) Rabbit mAb #86132:使用 SimpleChIP® Plus Enzymatic Chromatin IP Kit (Magnetic Beads) #9005 对 mES 细胞的交联染色质与 METTL3 (E3F2A) Rabbit mAb 或 Normal Rabbit IgG #2729 进行染色质免疫沉淀。使用 SimpleChIP® Mouse Sox2 Exon1 Primers #30180、mouse KLF2 intron 2 primers 和 SimpleChIP® Mouse MYT-1 Promoter Primers #8985 进行实时 PCR 来对富集的 DNA 进行定量。将每份样品中免疫沉淀的 DNA 的量表现为相对于所输入染色质总量(等于 1)的信号。
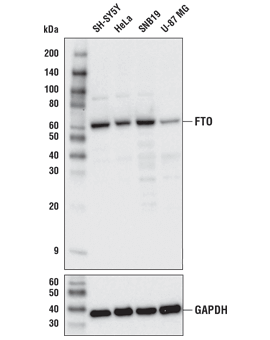
FTO (D6Z8W) Rabbit mAb #31687:使用 FTO (D6Z8W) Rabbit mAb(上图)或 GAPDH (D16H11) XP® Rabbit mAb #5174(下图)对不同细胞系的提取物进行蛋白印迹分析。
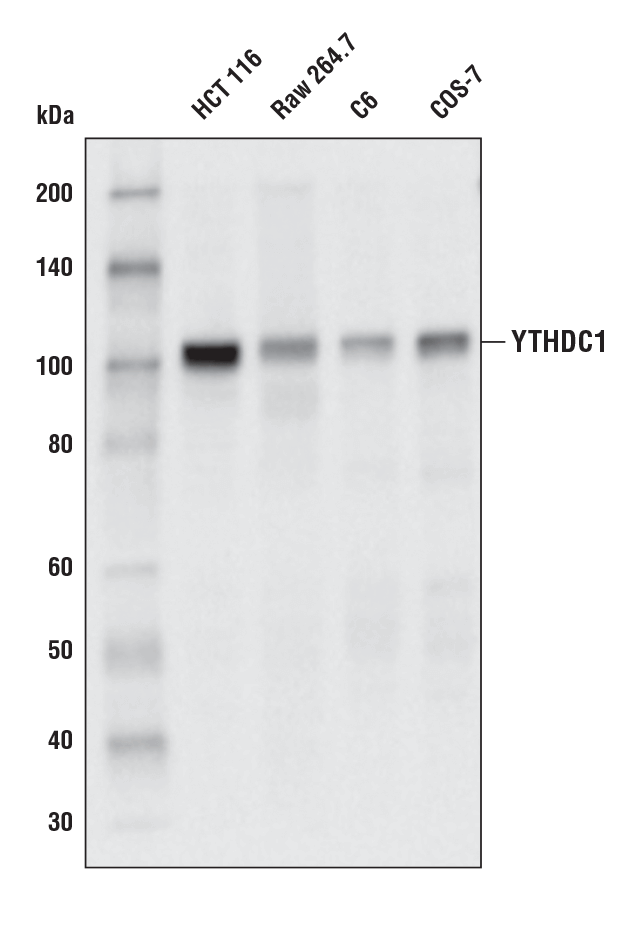
YTHDC1 (E4I9E) Rabbit mAb #77422:使用 YTHDC1 (E4I9E) 兔单克隆抗体对不同细胞系的提取物进行蛋白质印迹分析。
N6,2′-O-二甲基腺苷 (m6Am)
mRNA 在 5’ 端有 N7-甲基鸟苷 (m7G) 修饰,负责募集起始因子,并使细胞能够区分自我和病毒 mRNA (7)。当 m7G 帽之后的首个核苷酸 2′-O-甲基腺苷 (Am) 进一步被写入蛋白 CAPAM/PCIF1 甲基化形成 N6,2′-O-二甲基腺苷 (m6Am) 时,会发生可逆的甲基化修饰 (7)。通过 m6Am 擦除蛋白 FTO 即可去除该甲基。研究表明,CAPAM/PCIF1 甲基化促进了加帽 mRNA 的翻译,但其他研究观察到 m6A m 修饰 RNA 相较其 Am 修饰 RNA 的稳定性增加。
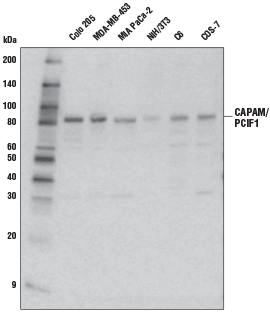
CAPAM/PCIF1 Antibody #98085:使用 CAPAM/PCIF1 Antibody 对不同细胞系的提取物进行蛋白印迹分析。
N1-甲基腺苷 (m1A)
不像 m6A 和 m6Am,N1 甲基腺苷 (m1A) 修饰会干扰 Watson-Crick 碱基配对,从而可能干扰蛋白-DNA 相互作用 (8)。它还被认为能调节 tRNA 和 rRNA 稳定性。与 m6A 和 m6Am 修饰相比,对于这种修饰明显知之甚少,因为检测到它有难度。mRNA 是否存在 m1A 修饰是不确定的,可能是因为使用的测序技术存在差异,因为有些团队检测到它比较富集,而其他团队压根没有检测到它。观察到 mRNA 存在 m1A 修饰的团队发现 ALKBH3 是一种 mRNA m1A 擦除蛋白,但并未检测到 m1A 写入蛋白 (9)。
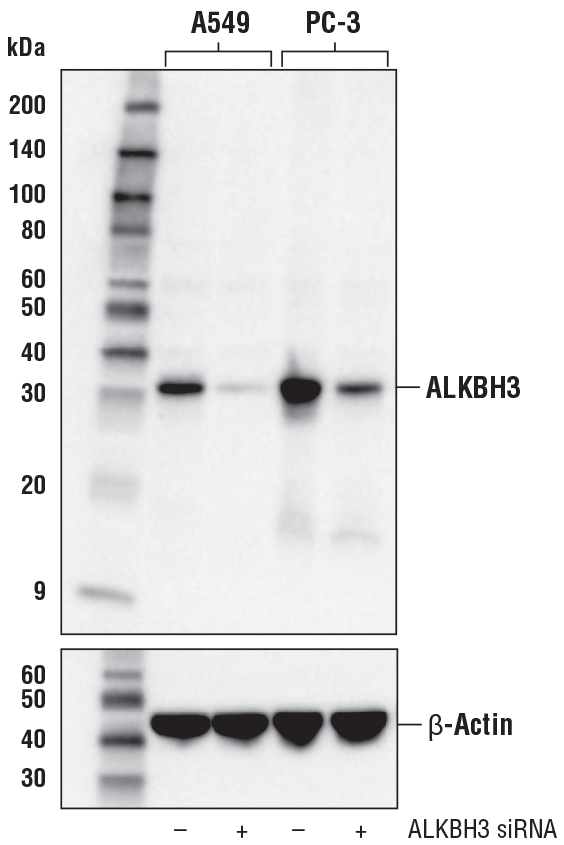
ALKBH3 (E6S4R) Rabbit mAb #87620: 使用 ALKBH3 (E6S4R) Rabbit mAb(上图)或 β-Actin (D6A8) Rabbit mAb #8457(下图)对转染空载 (-) 或转染 ALKBH3 siRNA (+) 的 A549 和 PC-3 细胞的提取物进行蛋白印迹分析。
其他 RNA 修饰
假尿苷
假尿苷是一种修饰,其中尿嘧啶中的氮碳键用碳-碳键替代。Dyskerin 等假尿苷合成酶(PUS 蛋白)主要在 tRNA 上催化这种变化,以促进 tRNA 稳定性 (10)。PUS 蛋白还可修饰 rRNA 以协助核糖体组装,这是正常蛋白合成所必需的。mRNA 假尿苷化最近才被观察到,并且可能影响 mRNA 剪切、翻译、半衰期和翻译调节。
Dyskerin 蛋白的异常活性与癌症进展和患者预后不良相关。假尿苷化还在 HIV 感染的病毒延迟过程调节中起作用,并且与克罗恩病、乳糜泻、X-连锁先天性角化不良 (X-DC)、母体遗传性糖尿病和耳聋 (MIDD)、线粒体肌病和铁粒幼细胞贫血 (MLASA) 有关。
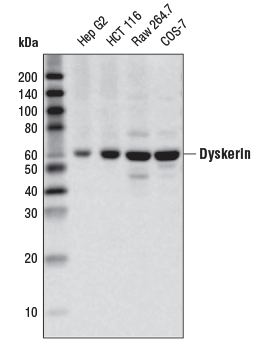
Dyskerin (D6N4K) Rabbit mAb #53234:使用 Dyskerin (D6N4K) Rabbit mAb 对不同细胞系提取物进行蛋白质印迹分析。
腺苷-肌苷转化
作用于 RNA 的腺苷脱氨酶 (ADAR) 会催化前 mRNA 中的腺苷脱氨形成肌苷。这种反应也称 A 至 I 编辑,它会改变 RNA 剪切和 RNA 成熟,导致蛋白编码和功能发生变化。此外,近期研究表明,A 至 I 编辑可防止对自我 RNA 产生天然免疫应答 (11)。ADAR1 编辑可以使双链 RNA (dsRNA) 变得不稳定,并抑制 RIG-I 或 MDA5 的激活,RIG-I 和 MDA5 是非自我物质识别和清除所涉及的两种监视蛋白。
通过改变 AZIN1 等蛋白的编码和行为,A 至 I 编辑会导致癌症进展。AZIN1 的 A 至 I 编辑导致丝氨酸在残基 367 被甘氨酸取代,增强了抑制多个肿瘤蛋白降解的能力,这可促进细胞运动、侵袭和转移。此外,癌细胞中的 ADAR1 丢失或敲除会克服对癌症免疫疗法产生抗性 (12)。
A 至 I 编辑似乎调节 HIV 感染,但目前尚未对其如何调节疾病达成共识。虽然 HIV-1 RNA 的 A 至 I 编辑已证实会中断病毒组装,并抑制原代巨噬细胞的增殖,但 ADAR1 活性增加与 CD4+ T 细胞中的 HIV-1 病毒复制增加相关 (11)。要了解 A 至 I 编辑对 HIV 感染的影响,还需更多的工作。
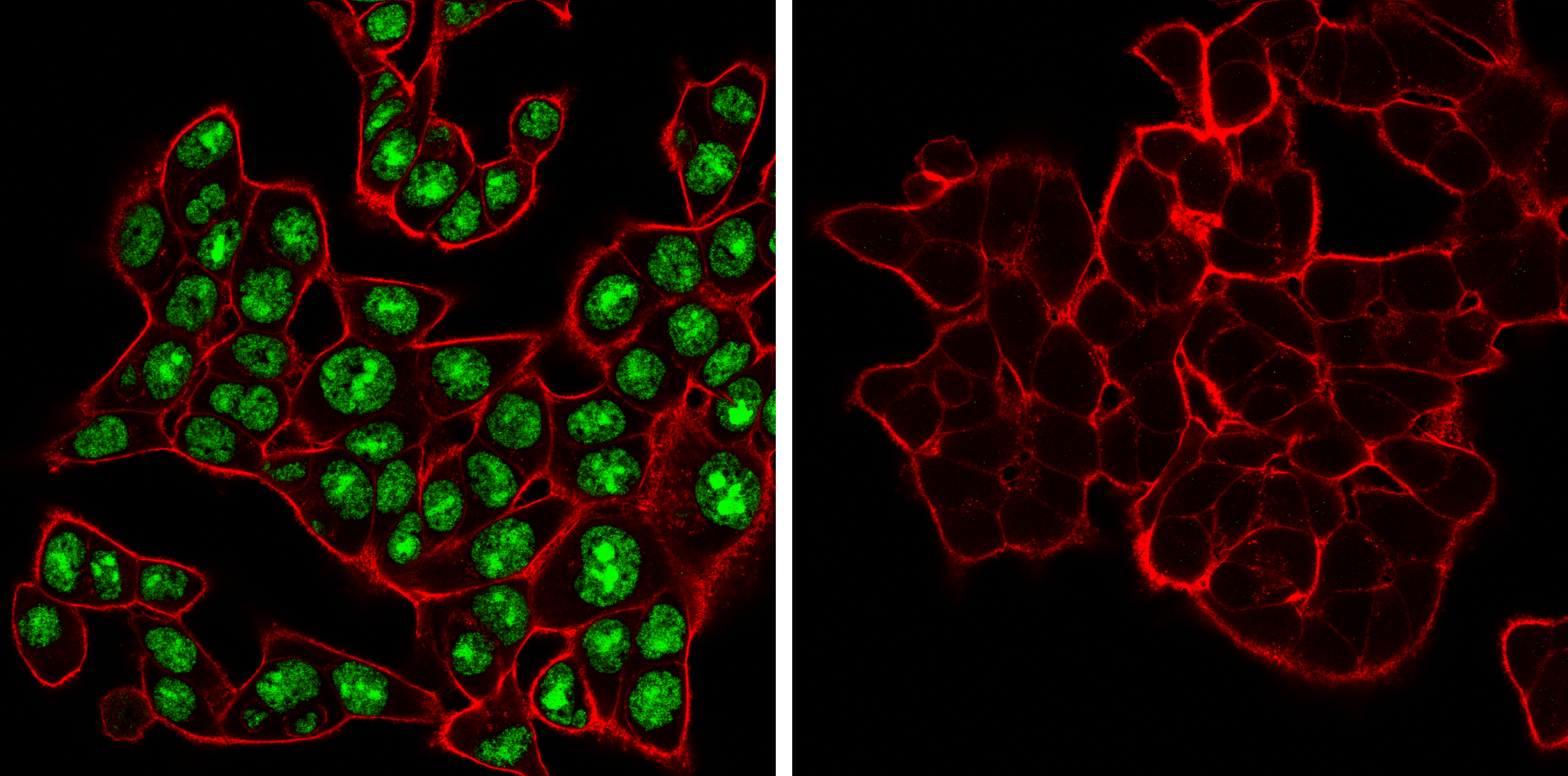
ADAR (E6X9R) XP® Rabbit mAb #81284: 使用 ADAR1 (E6X9R) XP® Rabbit mAb(绿色)对野生型(左图,阳性)或 ADAR1 敲除型(右图,阴性)HCT 116 细胞进行共聚焦免疫荧光分析。肌动蛋白丝用 DyLight 650 Phalloidin #12956(红色)标记。
选择性剪切
选择性剪切是一种受调节的过程,其中选择性 mRNA 转录物是从单个基因产生的,导致同一蛋白的不同同工型的翻译,这些同工型可以赋予不同的功能、结合伴侣和/或亚细胞定位。选择性剪切的基本模式包括组成型剪切、内含子保留、外显子跳过、选择性剪切位点选择、互斥剪切、选择性启动子选择,以及选择性多腺苷酸化位点 (13)。选择性剪切受蛋白 [例如 PTBP1 以及不均一核糖核蛋白(例如结合前 mRNA 且被剪切体催化的 hnRNP A1)]调控。
剪切体见于真核细胞的胞核,它是由小胞核 RNA (snRNA) 和蛋白组装而来的大分子组。主要剪切体是真核细胞中最富集的,并且包含含有 U1、U2、U4、U5 和 U6 snRNA 的 snRNP,但小剪切体不太常见,包含含有 U11、U12、U4atac、U4 和 U5 snRNA 的 snRNP (14)。剪切体可识别不同的序列元素,例如前 mRNA 中 5’ 端剪切位点,随后去除非编码内含子,并将侧翼外显子缝合在一起,以产生成熟的剪切 mRNA。核心剪切体还会募集 SAM68 等蛋白以协助选择性剪切。
SAM68 是一种调节 CD44 mRNA 和其他 RNA 的选择性剪切的 RBP。SAM68 及其选择性剪切靶标的许多活动与脊髓性肌肉萎缩症和癌症的发生有关 (15)。TDP-43 是一种调节 RNA 剪切、稳定性和运输的蛋白,在前颞叶变性和肌萎缩性脊髓侧索硬化症患者中,它会发生异常泛素化、磷酸化和裂解活化,从而产生会形成不溶聚合物的片段 (16)。最后,UAP56(ATP 依赖性剪切因子)还会促成与转移性去势抵抗前列腺癌相关的雄激素受体 AR-V7 剪切变体 (17, 18)。已知在癌症进展和/或治疗抗性中起到关键作用的其他剪切变体包括:与各种癌症有关的 12 种不同 p53 剪切变体 (19)。临床研究表明,p53 同工型的表达特征与肿瘤进展和临床效果相关。
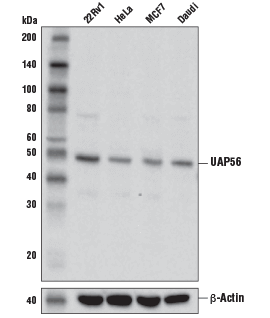
UAP56 Antibody #55509:使用 UAP56 抗体(上图)和 β-肌动蛋白 (D6A8) 兔单克隆抗体 #8457(下图)对不同细胞系的提取物进行蛋白印迹分析。
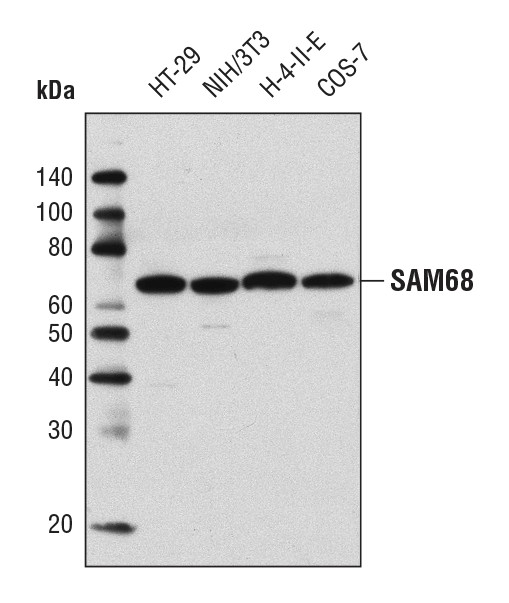
SAM68 Antibody #12538:使用 SAM68 Antibody 对不同细胞系的提取物进行蛋白质印迹分析。
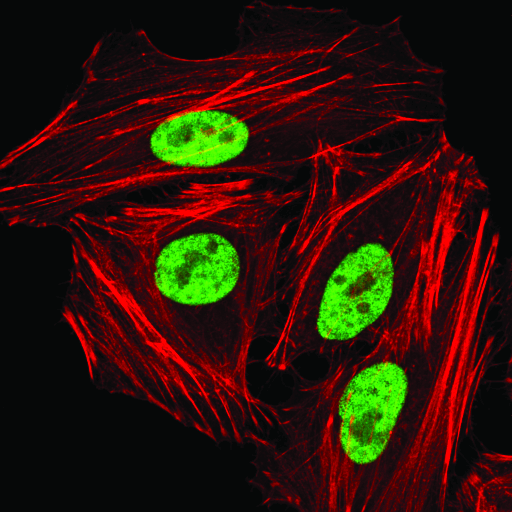
hnRNP A1 (D21H11) Rabbit mAb #8443:使用 hnRNP A1 (D21H1) Rabbit mAb(绿色),对 HeLa 细胞进行共聚焦免疫荧光分析。肌动蛋白丝用 DyLight 554 Phalloidin #13054(红色)标记。
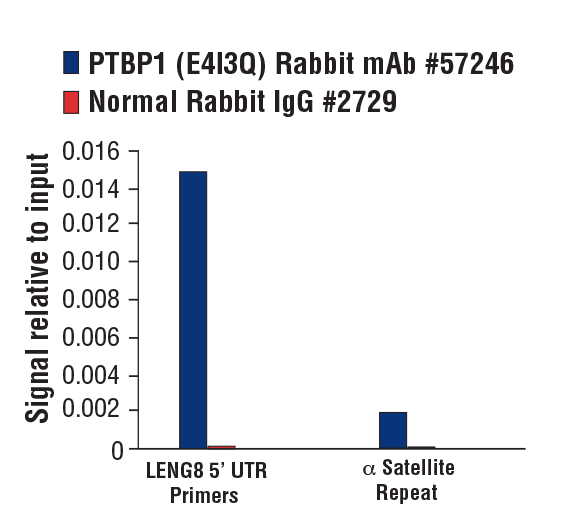
PTBP1 (E4I3Q) Rabbit mAb #57246:使用 SimpleChIP® 酶解染色质免疫共沉淀试剂盒(磁珠法) #9005 对在无酚红培养基和 5% 活性炭剥离的 FBS 中生长 4 天,随后用 β-雌二醇(10 nM,45 分钟)处理的 MCF7 细胞的交联染色质与 PTBP1 (E4I3Q) 兔单克隆抗体或正常兔 IgG #2729 进行染色质免疫沉淀。使用 SimpleChIP® Human LENG8 5’ UTR Primers #70823 和 SimpleChIP® Human α Satellite Repeat Primers #4486 进行实时 PCR 来对富集的 DNA 进行定量。将每份样品中免疫沉淀的 DNA 的量表现为相对于所输入染色质总量(等于 1)的信号。
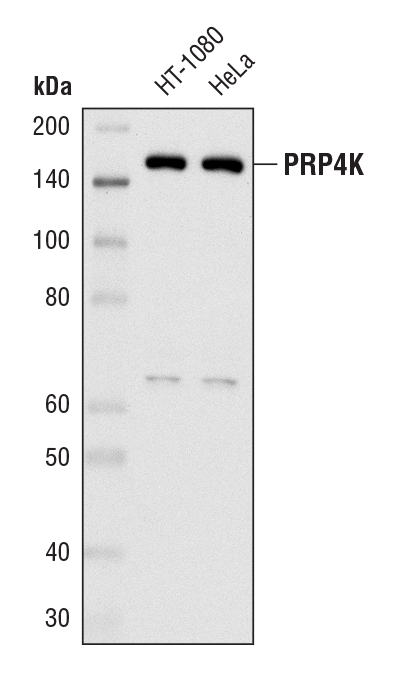
PRP4K (D27A1) Rabbit mAb #8577:使用 PRP4K (D27A1) Rabbit mAb 对 HT-1080 和 Hela 细胞的提取物进行蛋白质印迹分析。
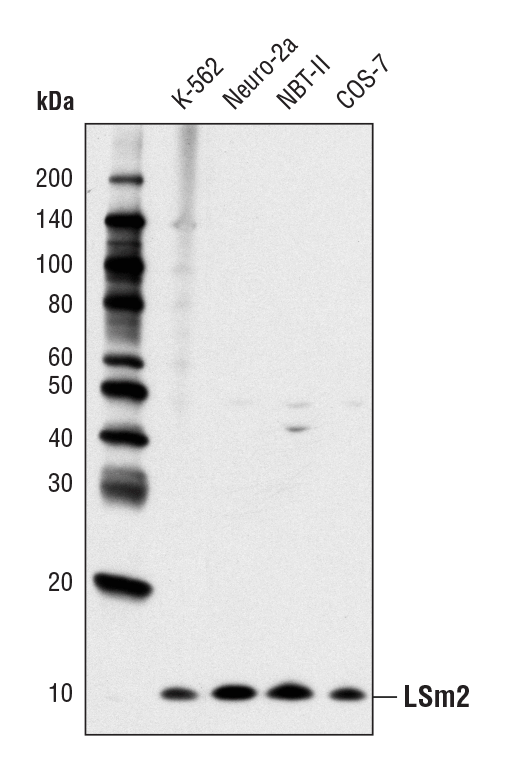
LSm2 (D9X6C) Rabbit mAb #13119:使用 LSm2 (D9X6C) Rabbit mAb 对不同细胞系的提取物进行蛋白质印迹分析。
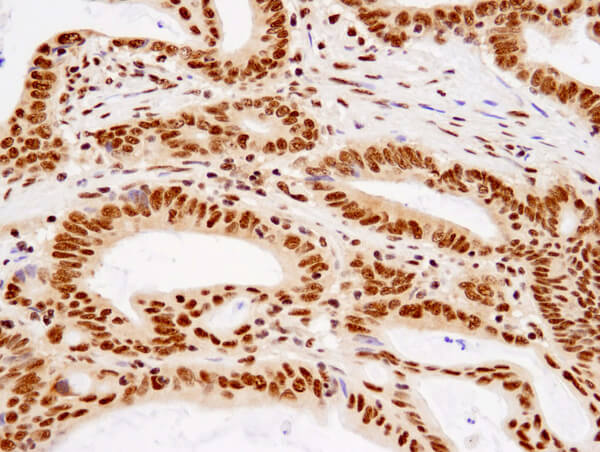
TDP43 (D9R3L) Rabbit mAb #89789:使用 TDP43 (D9R3L) Rabbit mAb 对石蜡包埋的人结肠癌细胞进行免疫组织化学分析
miRNA 分子组 (AGO/Dicer)
miRNA 是非编码 RNA,可降解或抑制靶标 mRNA 的表达,并调节转录后基因表达 (20)。它们被 RNA 聚合酶 II 转录为初级 miRNA (pri-mRNA),然后在胞核中被 Drosha 和 DGCR8 裂解形成前 miRNA。这些前 miRNA 包含短发夹,随后被 XPO5 导出到细胞浆,并在其中被 Dicer 进一步加工成成熟的 miRNA。成熟 miRNA 接着与 Argonaute 2 (AGO2) 和 TRBP 等蛋白相互作用,形成 RNA 诱导的沉默复合体 (RISC)。RISC 依赖性基因沉默在 mRNA 降解或翻译抑制时发生。
miRNA 分子组扰动,例如 AGO2、Drosha 和 DGCR8 表达改变,经证实会在很多疾病条件下导致癌变。miRNA 失调还与心脏和肾脏疾病有关 (21, 22)。
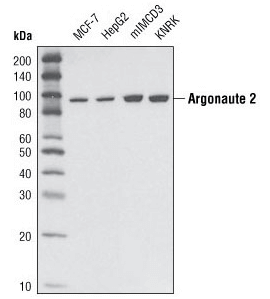
Argonaute 2 (C34C6) Rabbit mAb #2897:使用 Argonaute 2 (C34C6) Rabbit mAb 对不同细胞类型的提取物进行蛋白质印迹分析。
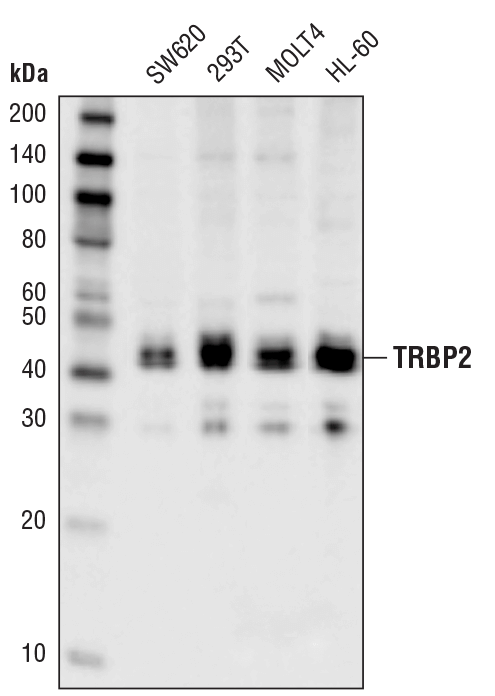
TRBP2 (D7C8K) Rabbit mAb #62043:使用 TRBP2 (D7C8K) Rabbit mAb 对来自不同细胞系的提取物进行蛋白质印迹分析。
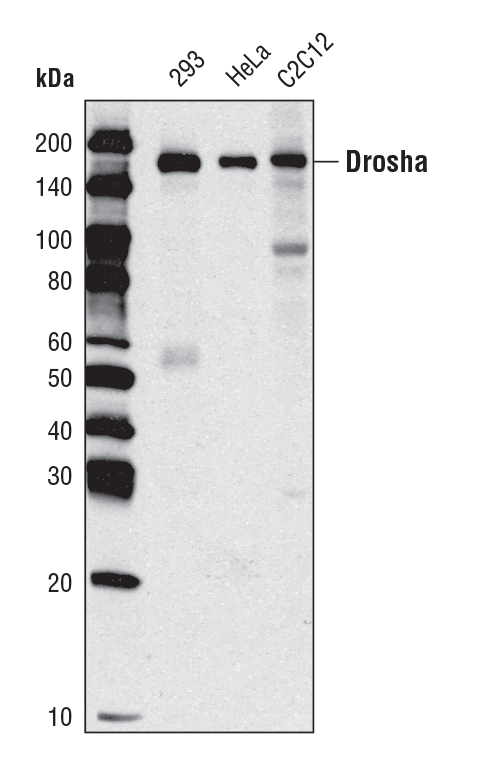
Drosha (D28B1) Rabbit mAb #3364:使用 Drosha (D 28B1) Rabbit mAb 对不同细胞类型的提取物进行蛋白印迹分析。

Dicer (D38E7) Rabbit mAb #5362:使用 Dicer (D38E7) Rabbit mAb 对 MCF7 和 293 细胞提取物进行蛋白质印迹分析。
RNP 颗粒
mRNA 与 RBP 相互作用形成 mRNP 颗粒,mRNP 颗粒会调节 mRNA 定位、稳定性和翻译。在许多不同类型的颗粒中,应激颗粒和处理体的特征最明显 (23)。处理体是含有脱帽酶、翻译调节分子、miRNA 分子组以及与无义介导的衰变有关的因子的细胞浆 RNA 颗粒。应激颗粒可在细胞应激过程中保护特定 mRNA,并且包含延伸因子、40 核糖体亚基和停滞 mRNA。它们在应激诱导的 eIF2 磷酸化后形成。G3BP1(一种与 RasGAP 的 SH3 结构域相互作用的蛋白)常用作应激颗粒标记物,因为它对应激颗粒的组装至关重要,并且是一种应激颗粒成核蛋白 (24)。
应激颗粒的异常或持续形成会导致肌萎缩性脊髓侧索硬化症、佩吉特氏病和额颞叶变性等退行性疾病 (25)。有这些疾病状态的细胞含有有许多与应激颗粒相同的组分的包涵体。另外,许多与这些疾病有关的突变蛋白都见于应激颗粒,或导致异常应激颗粒形成的过程中。有证据还表明应激颗粒与癌症之间存在关系,但导致疾病进展的机制尚不清楚 (23)。
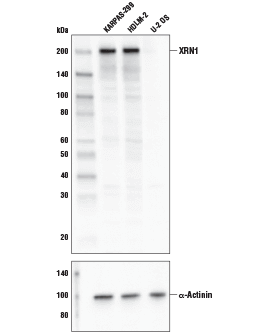
XRN1 Antibody #70205:使用 XRN1 Antibody(上图)和 α-Actinin (D6F6) XP® Rabbit mAb #6487(下图)对 KARPAS-299、HDLM-2 和 U-2 OS 细胞系的提取物进行蛋白质印迹分析。KARPAS 细胞系来源:剑桥大学 Abraham Karpas 博士。
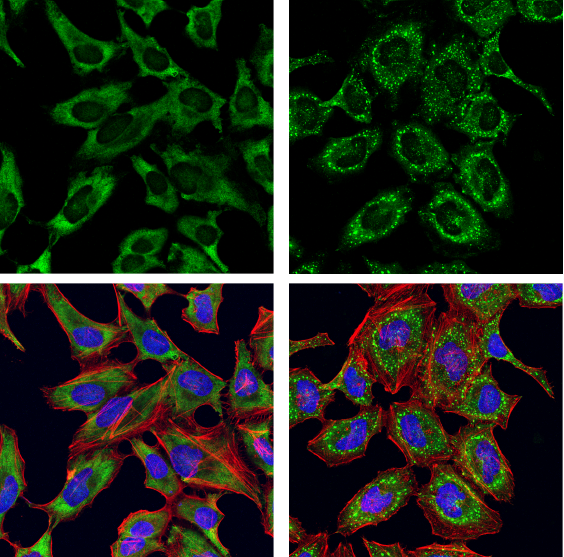
G3BP1 Antibody #17798:使用 G3BP1 抗体(绿色)对血清饥饿(左图)或血清饥饿后经亚砷酸钠(500 μm,30 分钟;右图)处理的 HeLa 细胞进行共聚焦免疫荧光分析。肌动蛋白丝用 DyLight 554 Phalloidin #13054(红色)标记。样品用 ProLong Gold Antifade Reagent with DAPI #8961(蓝色)封片。
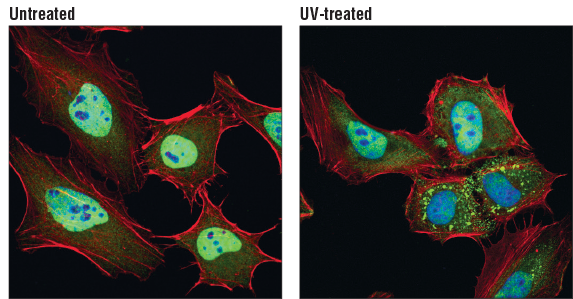
TIAR (D32D3) XP® Rabbit mAb #8509:使用 TIAR (D32D3) XP® Rabbit mAb(绿色),对未经处理的(左图)或经紫外线处理的(右图) HeLa 细胞进行共聚焦免疫荧光分析。肌动蛋白丝用 DyLight 554 Phalloidin #13054(红色)标记。蓝色伪彩 = DRAQ5 #4084(DNA 荧光染料)。
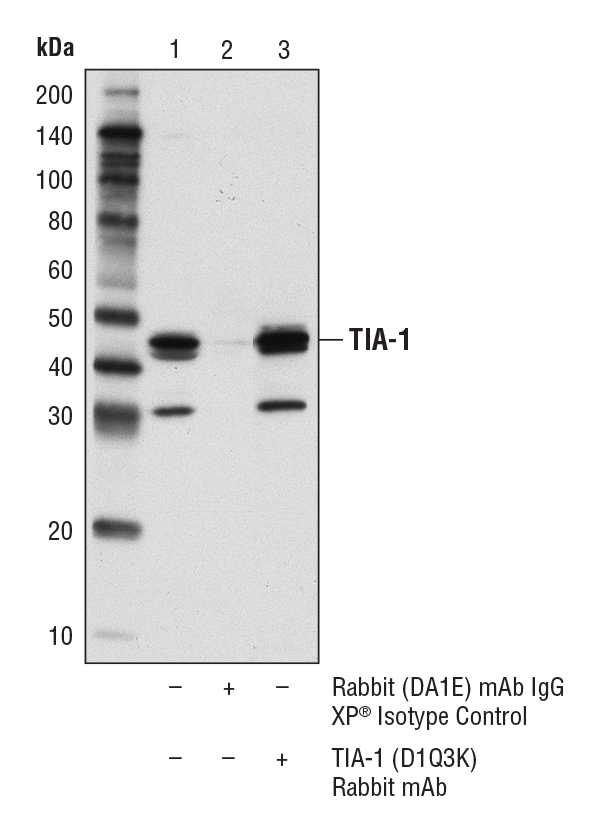
TIA-1 (D1Q3K) Rabbit mAb #86050:对 KARPAS-299 细胞裂解物进行 TIA-1 的免疫沉淀。泳道 1 为 10% input,泳道 2 为 Rabbit (DA1E) mAb IgG XP® Isotype Control #3900,泳道 3 为 TIA-1 (D1Q3K) Rabbit mAb。使用 TIA-1 (D1Q3K) Rabbit mAb 进行蛋白质印迹。使用具有构象特异性二抗,来避免与 IgG 发生交叉反应。细胞系来源:剑桥大学 Abraham Karpas 博士。
参考文献
- Kapeli K and Yeo GW. (2012) Front Neurosci 6:144
- Geuens T, Bouhy D, Timmerman V. (2016) Hum Genet 135(8):851-867
- Wang S, et al. (2018) Mol Cancer 17(1):101
- Luo J, et al. (2018) Int J Mol Sci 19(9)
- Chen X, et al. (2019) ACS Chem Neurosci 10(5):2355-2363
- Reitz C, et al. (2012) PLoS One 7(12):e50354
- Mauer J, et al. (2017) Nature 541:371-375
- Akichika S, et al. (2019) Science 363(6423)
- Zhang C and Jia G. (2018) Genomics, Proteomics & Bioinformatics 16(3):155-161
- Penzo M, et al. (2017) Genes (Basel) 8(11)
- Song C, et al. (2016) Genes (Basel) 7(12)
- Ishizuka JJ, et al. (2019) Nature 565(7737):43-48
- Wang BD and Lee NH. (2018) Cancers (Basel) 10(11):pii. e458
- Chen W and Moore MJ. (2015) Curr Biol 5(5):R181-3
- Matter N, et al. (2002) Nature 420:691-695
- Jeon GS, et al. (2019) Mol Neurobiol 56(3):2007-2021
- Nakata D, et al. (2017) Biochem Biophys Res Commun 483:271-276
- Xu J and Qiu Y., (2016) Asian J Urol 3(4):177-184
- Surget S, Khoury MP, and Vourdon JC. (2013) Onco Targets Ther 7:57-68
- Huang JT, et al. (2014) Front Oncol 4:113
- Tatsuguchi M, et al. (2007) J Mol Cell Cardiol 42(6):1137-1141
- Phua YL, et al. (2015) Physiol Rep 3(10):pii. e12537
- Kedersha N, et al. (2016) J Cell Biol 212:845-60
- Andrea P, Kedersha N, and Ivanov P. (2015) Biochim Biophys Acta 1849(7):861-870
- Buchan JR. (2014) RNA Biol 11(8):1019-1030